브로카의 면적
Broca's area| 브로카의 면적 | |
|---|---|
 Broca의 영역은 Brodmann 영역 44(pars opercularis)와 45(pars triangularis)로 구성되어 있습니다. | |
 Broca의 영역(빨간색으로 표시) | |
| 세부 사항 | |
| 의 일부 | 전두엽 |
| 동맥 | 중뇌 |
| 정맥 | 상악동 |
| 식별자 | |
| MeSH | D065711 |
| 뉴로네임즈 | 2062 |
| FMA | 242176 |
| 신경해부학의 해부학적 용어 [위키데이터 편집] | |
Broca's area, or the Broca area (/ˈbroʊkə/,[1][2][3] also UK: /ˈbrɒkə/, US: /ˈbroʊkɑː/[4]), is a region in the frontal lobe of the dominant hemisphere, usually the left, of the brain[5] with functions linked to speech production.
언어 처리는 Pierre Paul Broca가 두 명의 환자에게서 장애를 보고한 이후로 Broca의 영역과 연결되어 왔습니다.[6] 그들은 뇌의 후방 하전두회 (pars triangularis) (BA45)에 부상을 입은 후에 말하는 능력을 잃었습니다.[7] 이후 그가 파악한 대략적인 지역은 브로카의 지역으로 알려지게 되었고 언어 생산의 적자는 표현성 실어증이라고도 불리는 브로카의 실어증으로 알려지게 되었습니다. Broca의 영역은 이제 일반적으로 하전두회의 pars opercularis와 pars triangularis로 정의되며, Brodmann의 세포건축학 지도에서 Brodmann 영역 44와 Brodmann 영역 45로 표시됩니다.[7]
기능적 자기 공명 영상(fMRI)은 언어 처리가 BA6의 복부 부분뿐만 아니라 하전두회의 세 번째 부분을 포함하는 것으로 나타났으며, 이들은 종종 브로카 영역이라고 불리는 더 큰 영역에 포함됩니다.[8]
만성 실어증에 대한 연구는 다양한 언어 및 언어 기능에서 브로카 영역의 필수적인 역할을 암시했습니다. 또한 fMRI 연구를 통해 다양한 언어 작업과 관련된 Broca 영역의 활성화 패턴도 확인되었습니다. 그러나 뇌종양에 의한 브로카 영역의 느린 파괴는 언어를 비교적 손상시키지 않을 수 있으며, 이는 그 기능이 뇌의 가까운 영역으로 이동할 수 있음을 시사합니다.[9]
구조.


브로카의 영역은 종종 설시와 같은 거대 구조적 랜드마크나 특정 기준 공간의 좌표 지정에 의해 뇌의 지형을 시각적으로 검사하여 식별됩니다. 현재 사용되는 Talairach와 Tournoux 지도는 Brodmann의 세포 건축학 지도를 템플릿 뇌에 투영합니다. Brodmann의 파세레이션은 세포 건축학적 경계에 대한 주관적인 시각적 검사를 기반으로 했고 Brodmann 또한 한 뇌의 한 반구만을 분석했기 때문에 결과는 부정확합니다. 또한, 설골 및 자이랄 구조에 비해 형태, 크기 및 위치 측면에서 뇌 전반에 걸쳐 상당한 가변성 때문에 결과적인 국소화 정밀도가 제한됩니다.[10]
그럼에도 불구하고, 좌반구의 브로카 영역과 우반구의 상동체는 일반적으로 하전두회(PTR)의 삼각 부분과 하전두회(POP)의 오큘러 부분을 지칭하는 데 사용되는 명칭입니다. PTr 및 POP는 Brodmann의 분류 체계에 의해 하전두회를 각각 45 및 44의 전방 및 후방 세포 건축 영역으로 확률적으로만 분할하는 구조적 랜드마크에 의해 정의됩니다.[11]
영역 45는 운동, 체성감각 및 하대정골 영역으로부터 더 많은 구심성 연결을 받는 경향이 있는 영역 44에 비해 전전두엽 피질, 상측두엽 및 상측두엽으로부터 더 많은 구심성 연결을 받습니다.[11]
세포 아키텍처 및 연결성에서 영역 45와 44의 차이점은 이러한 영역이 다른 기능을 수행할 수 있음을 시사합니다. 실제로 최근 신경영상 연구에 따르면 각각 45번과 44번 영역에 해당하는 PTr과 Pop은 언어 이해 및 행동 인식/이해와 관련하여 인간에서 다른 기능적 역할을 수행합니다.[11]
브로카의 면적은 남성보다 여성이 약 20% 더 큽니다.[12]
기능들
언어 이해력
오랫동안 브로카의 영역의 역할은 언어 이해보다 언어 생산에 더 헌신적이라고 가정했습니다. 그러나 브로카의 영역이 언어 이해에도 중요한 역할을 한다는 것을 증명하는 증거가 있습니다. 문법적 음성 연출을 보이는 Broca 영역의 병변이 있는 환자들도 구문 정보를 사용하여 문장의 의미를 결정할 수 없음을 보여줍니다.[13] 또한, 많은 신경 영상 연구에서 복잡한 문장을 처리하는 동안 브로카 부위, 특히 왼쪽 하전두회의 파스 오큘라리스가 관련되어 있습니다.[14] 또한 기능적 자기공명영상(fMRI) 실험은 매우 모호한 문장이 더 활성화된 하전두회를 초래한다는 것을 보여주었습니다.[15] 따라서, 매우 모호한 내용과 관련된 검색 요구가 증가하기 때문에, 하전두회의 활동 수준과 어휘 모호성 수준은 서로 정비례합니다.
또한 Broca의 영역 내에서 이해의 특정 측면에 대한 전문성이 있습니다. Devlin 등에 의한 작업. (2003)[16]은 반복적 경두개 자기 자극(rTMS) 연구에서 pars triangularis(Broca 영역의 전방에 위치)를 목표로 하는 rTMS 하에서 의미 과제를 수행할 때 반응 시간이 증가하는 것으로 나타났습니다. 반응 시간의 증가는 특정 영역이 그 인지 기능을 처리하는 역할을 한다는 것을 나타냅니다. TMS를 통해 이러한 영역을 방해하면 해당 영역에서 수행되는 계산이 방해되어 계산을 수행하는 데 필요한 시간이 증가합니다(반응 시간에 반영됨). 닉슨 등의 후작. (2004)[17]는 pars opercularis (Broca 영역의 뒤쪽에 위치함)가 rTMS 하에서 자극되었을 때 음운학적 과제에서 반응 시간이 증가했음을 보여주었습니다. Gough et al. (2005)[18]는 Broca 영역의 전방 또는 후방을 향하는 rTMS 자극으로 음운적 작업과 의미적 작업이 모두 수행되는 이러한 이전 작업의 요소를 결합하는 실험을 수행했습니다. 이 실험의 결과는 언어 이해의 다양한 구성 요소에 대해 Broca의 영역 내 해부학적 전문성을 결정적으로 구별했습니다. 여기서 결과는 rTMS 자극 하에서 다음과 같이 나타났습니다.
- 의미론적 과제는 자극이 Broca 영역의 전방을 목표로 했을 때 반응 시간이 감소하는 것만 나타냈습니다(비 TMS 대조군에 비해 10%(50 ms)의 감소가 관찰됨).
- 음운학적 과제는 Broca 영역의 뒤쪽을 자극 대상으로 했을 때 반응 시간이 감소하는 것으로 나타났습니다(대조군에 비해 6%(30 ms)의 감소가 관찰되었습니다).
요약하자면, 위의 작업은 언어 이해를 위한 브로카 영역의 해부학적 전문성을 보여주며, 브로카 영역의 앞부분은 단어의 의미를 이해하는 역할을 하고, 뒷부분은 단어가 어떻게 소리가 나는지를 이해하는 역할을 합니다.
동작인식 및 제작
최근의 실험들은 브로카의 영역이 다양한 인지 및 지각 작업에 관여한다는 것을 나타냈습니다. Brodmann의 영역 44의 한 가지 중요한 기여는 모터 관련 프로세스에서도 발견됩니다. 움직이는 동물을 닮은 의미 있는 손 그림자를 관찰하면 정면 언어 영역이 활성화되어 브로카의 영역이 실제로 다른 사람의 행동을 해석하는 역할을 한다는 것을 보여줍니다.[19] 파지 및 조작을 실행하는 동안 BA 44의 활성화도 보고되었습니다.[20]
음성 관련 제스처
음성 관련 제스처가 어휘적 또는 본질적 모호성을 감소시킬 수 있기 때문에 음성 관련 제스처가 있는 경우 이해력이 향상되어야 한다고 추측되어 왔습니다. 이해력이 향상된 결과로 브로카 영역의 참여를 줄여야 합니다.[11]
많은 신경 영상 연구에서 의미 있는 팔 동작을 나타낼 때 브로카 부위의 활성화도 보여주었습니다. 최근의 연구는 단어와 제스처가 운동 목표와 의도와 같은 특정 제스처 측면의 번역 수준에서 관련이 있다는 증거를 보여주었습니다.[21] 이 발견은 왜 이 부분에 결함이 있을 때 수화를 사용하는 사람들도 언어 결손이 있는지를 설명하는 데 도움이 됩니다.[22]: 494–7 제스처의 측면이 브로카의 영역 내에서 말로 번역된다는 이 발견은 언어 발달을 진화의 측면에서도 설명합니다. 실제로, 많은 저자들은 말이 몸짓에서 비롯된 원시적인 의사소통에서 진화했다고 제안했습니다.[19][23] (아래 참조).
브로카 영역 없이 말하기
Broca 영역의 손상은 일반적으로 내용 어휘로 구성된 전신 음성과 관련이 있습니다. 예를 들어, 브로카 실어증을 앓고 있는 사람은 이렇게 말할지도 모릅니다. "운전, 가게. 엄마"라는 말은 "엄마가 오늘 가게까지 데려다줬어요"라는 뜻입니다. 따라서 정보의 내용은 맞지만 문장의 문법과 유동성이 빠져 있습니다.[24]
언어를 거의 그대로 둔 채 파괴할 수 있다는 점에서 브로카 지역의 음성 제작에서의 본질적인 역할에 의문이 제기되고 있습니다. 컴퓨터 엔지니어의 한 사례에서 느리게 성장하는 교종 종양을 제거했습니다. 종양과 수술로 왼쪽 하반신과 중간 전두회, 미상핵의 머리, 내부 캡슐의 전지, 그리고 전방 절연체가 파괴되었습니다. 하지만 제거 후 3개월이 지나 언어 문제가 최소한으로 발생했고 개인은 전문적인 업무로 복귀했습니다. 이런 사소한 문제들은 두 개 이상의 주제를 포함한 구문론적으로 복잡한 문장을 만들 수 없는 것, 복수의 인과 관계나 보고된 음성을 포함합니다. 이것들은 연구자들에 의해 작업 기억 문제 때문이라고 설명되었습니다. 그들은 또한 그의 문제가 없는 것은 인근 대뇌 피질의 신경 가소성과 우반구의 상동 영역으로의 일부 기능 이동에 의해 가능해진 광범위한 보상 메커니즘 때문이라고 설명했습니다.[9]
임상적 의의
버벅버벅
말더듬이라고 알려진 언어 장애는 브로카의 부위의 활동 부족과 관련이 있는 것으로 보입니다.[25][26]
아파시아
실어증은 쓰기, 읽기, 말하기, 듣기 등 모든 양식에 영향을 미치는 후천적 언어 장애로 뇌 손상에서 비롯됩니다. 자신의 삶의 모든 영역에 변화를 일으키는 만성 질환인 경우가 많습니다.[27]
발현성 실어증 대 기타 실어증
브로카 실어증이라고도 불리는 표현성 실어증 환자들은 "그들이 하고 싶은 말, 그들은 단지 그것을 꺼낼 수 없다"는 것을 알고 있는 개인들입니다.[27] 그들은 일반적으로 단어와 문장을 이해할 수 있지만(위 참조), 유창한 발화를 만들어낼 수는 거의 없습니다. 다른 증상으로는 유창성, 발화, 단어 찾기, 단어 반복, 복잡한 문법 문장의 생성과 이해에 대한 문제가 있을 수 있습니다.[7]
이 특정 증상 그룹은 표현성 실어증을 가진 사람과 다른 유형의 실어증을 가진 사람을 구별합니다. 실어증에는 몇 가지 뚜렷한 "유형"이 있으며, 각 유형은 다른 언어 결핍을 특징으로 합니다. 표현성 실어증을 가진 사람들은 좋은 언어 이해력을 유지하는 경향이 있지만, 다른 유형의 실어증은 환자가 어떤 언어도 완전히 이해할 수 없게 만들고, 어떤 언어도 이해할 수 없게 만들 수 있지만(청각성 언어 불가지증),[28][29][30] 다른 유형은 언어 이해력을 유지하지만 결함이 있습니다. 표현성 실어증을 가진 사람들은 다른 종류의 실어증을 가진 사람들보다 읽기와 쓰기(알렉시아 참조)에 덜 어려움을 겪을 수 있습니다.[22]: 480–500 비록 표현성 실어증을 가진 사람들은 그들의 언어 출력을 스스로 모니터링하는 좋은 능력을 가지고 있는 경향이 있지만(그들은 그들이 말하는 것을 듣고 수정합니다), 다른 종류의 실어증들은 그들의 언어 결핍을 전혀 모르는 것처럼 보일 수 있습니다.
고전적인 의미에서 발현성 실어증은 브로카 부위의 손상의 결과이며,[31] 비록 사례 연구에 따르면 병변 위치와 실어증 증상 사이에 항상 일대일 매핑이 있는 것은 아니지만 특정 뇌 부위의 병변이 특정한 해리 가능한 증상을 일으키는 경우가 많습니다.[29] 특정한 뇌부위(보통 좌반구)의 손상과 특정한 종류의 실어증의 발생 사이의 상관관계는 특정한 종류의 실어증의 존재(및 중증도)만을 근거로 뇌병변이 의심되는 위치를 추론할 수 있게 하고, 이것은 환자가 여러 뇌 부위에 손상을 입었을 수 있고 한 종류 이상의 실어증 증상을 보일 수 있기 때문에 복잡합니다. 인지의 특정 측면의 정상적인 기능에 필수적인 뇌 영역을 추론하기 위해 병변 데이터를 조사하는 것을 결손-병변 방법이라고 하는데, 이 방법은 특히 신경과학의 한 분야에서 중요합니다. 인지 과학(구체적으로 인지 신경 심리학)은 결손-병변 방법을 광범위하게 사용하는 신경 과학의 한 분야입니다.[32]
| 실어증의 종류 | 음성반복 | 네이밍 | 청각 이해력 | 유창성 |
|---|---|---|---|---|
| 표현성 실어증 | 보통-심각 | 보통-심각 | 경도난도 | 유창하지 않고, 노력적이며, 느림 |
| 수용성 실어증 | 경증-심증 | 경증-심증 | 결함이 있는 | 유창한 의역 |
| 전도성 실어증 | 불쌍한. | 불쌍한. | 비교적 좋음 | 유창한 |
| 혼합 경피적 실어증 | 적당한. | 불쌍한. | 불쌍한. | 비유동적 |
| 경피적 운동성 실어증 | 좋아요. | 경증-심증 | 마일드 | 비유동적 |
| 경피적 감각 실어증 | 좋아요. | 보통-심각 | 불쌍한. | 유창한 |
| 전지구적 실어증 | 불쌍한. | 불쌍한. | 불쌍한. | 비유동적 |
| 아노믹 실어증 | 마일드 | 보통-심각 | 마일드 | 유창한 |
1970년대[33] 후반에 연구가 수행된 이래로 브로카의 지역과 브로카 실어증 사이의 관계는 생각했던 것만큼 일치하지 않는다는 것이 알려져 왔습니다.[34] 브로카 부위의 병변만으로는 브로카의 실어증이 발생하지 않으며, 브로카의 실어증 환자가 브로카 부위에 반드시 병변이 있는 것도 아닙니다.[35] Broca 부위의 병변만으로도 3-6주 이내에 해결되는 일시적인 돌연변이가 발생하는 것으로 알려져 있습니다. 이 발견은 브로카의 영역이 언어화 또는 발화의 어떤 측면에 포함될 수 있음을 시사합니다. 그러나 이것은 문장 이해에서 브로카의 영역을 다루지 않습니다. 그러나 문장 처리의 기능적 영상화 연구에서는 브로카의 영역이 자주 등장합니다.[36] 그러나 단어 수준 작업에서도 활성화됩니다.[37] 이는 Broca의 영역이 문장 처리에만 전념하는 것이 아니라 양자에 공통되는 기능을 지원하고 있음을 시사합니다. 사실 브로카의 영역은 움직임의 이미지와 같은 비언어적 작업에서 활성화를 보여줄 수 있습니다.[38]
브로카 영역이 아티큘레이션에 가장 관여할 수 있다는 가설을 고려할 때, 이 모든 작업에서 브로카 영역의 활성화는 응답을 공식화하는 동안 피실험자의 은밀한 아티큘레이션 때문일 수 있습니다. 이러한 주의 사항에도 불구하고, 브로카의 영역이 어떤 역할을 하든지 간에, 그것은 전두엽 영역의 알려진 작동 기억 기능과 관련이 있을 수 있다는 공감대가 형성되고 있는 것 같습니다. (Broca 영역의 일부로 언급되는 기능성 영상 문헌에 보고된 탈라라크[39] 좌표는 광범위하게 분포되어 있습니다.) 예를 들어, 수동 음성 문장의 처리는 문장의 다른 관련 부분이 조작되는 동안 정보의 일시적인 보존을 돕기 위해 작업 메모리를 필요로 할 수 있습니다(즉, 논쟁에 대한 주제적 역할의 할당을 해결하기 위해). 미야케, 카펜터, 저스트는 문장 처리가 이러한 일반적인 언어 작업 기억 메커니즘에 의존한다고 제안한 반면, 카플란과 워터스는 브로카의 영역이 구문 처리를 위해 작업 기억에 관여한다고 생각합니다. Friederici(2002)는 Broca의 영역을 구성 영역으로 구분하고 Brodmann의 영역 44가 음운적[40] 구조와 구문적 구조 모두에 대한 작업 기억에 관여한다고 제안합니다. 이 영역은 이해 과정의 시간 과정이 전개됨에 따라 음운학의 경우 먼저 활성화되고 구문의 경우 나중에 활성화됩니다. Brodmann의 영역 45와 Brodmann의 영역 47은 통사적 재분석과 수리의 과정이 요구되는 주제 구조와 의미적 특징을 위한 작업 기억에 구체적으로 관여하는 것으로 간주됩니다. 이 영역들은 Brodmann의 영역 44가 처리 역할을 마친 후 온라인에 접속되며 복잡한 문장의 이해가 일반적인 기억 자원에 의존해야 할 때 활성화됩니다. 이 모든 이론들은 통사적 이해 문제가 개념적인 결함이 아니라 계산적인 것에서 발생한다는 견해를 향한 움직임을 나타냅니다. 새로운 이론들은 뇌가 어떻게 다른 언어적 그리고 인지적 요소들을 통합하는지에 대한 더 역동적인 관점을 가지고 이러한 수술들의 시간 경과를 조사하고 있습니다.
신경 인지 연구는 이미 브로카의 영역에 인접한 전두엽 영역을 비언어적 작업뿐만 아니라 작업 기억에 중요하게 포함시켰습니다.[41] Cabeza와 Nyberg의 작업 메모리 영상 연구 분석은 BA45/47이 정보를 선택하거나 비교하기 위해 모집되는 반면, BA9/46은 작업 메모리의 정보 조작에 더 관여할 수 있다는 견해를 뒷받침합니다. Broca의 실어증을 생성하기 위해서는 일반적으로 큰 병변이 필요하기 때문에 이러한 영역은 일부 환자에서도 손상될 수 있으며 복잡한 형태 합성 구조에 대한 이해 부족에 기여할 수 있습니다.
Broca의 영역은 음운 순서의 연결에 있어서 핵심적인 중심지입니다.
브로카의 영역은 이전에 음운 분할, 통사적 처리, 통일 등 다양한 과정과 관련이 있었는데, 이 모든 과정은 다양한 유형의 언어 정보를 분할하고 연결하는 것을 포함합니다.[42][43][44] 단일 단어를 반복하고 읽는 것은 의미론적 및 구문론적 처리를 수반하지 않지만 음소 시퀀스와 운동 제스처를 연결하는 작업이 필요합니다. 연구 결과에 따르면 이 연결은 실제 발화 행위 이전에 운동 피질과의 상호 작용을 포함하여 음소 및 조음 표현을 각각 담당하는 시간 및 전두엽 피질과의 상호 작용을 통해 브로카의 영역에 의해 조정됩니다. 이러한 독특한 연구 결과를 바탕으로 브로카의 영역은 관절의 위치가 아니라 음성 생성의 주요 구성 요소를 담당하는 대규모 피질 네트워크에 걸쳐 신경 정보를 조작하고 전달하는 핵심 노드임이 제안되었습니다[by whom?].[citation needed]
역사
2007년 발표된 연구에서 Leborgne과 Leong(브로카 환자)의 보존된 뇌를 고해상도 체적 MRI를 사용하여 재검사했습니다. 이 연구의 목적은 뇌를 3차원으로 스캔하고 피질하 병변과 피질하 병변의 범위를 더 자세히 확인하는 것이었습니다. 이 연구는 또한 피질하 침범 범위를 가진 현재 브로카 영역이라고 불리는 것과 관련하여 전두엽에 있는 병변의 정확한 위치를 찾고자 했습니다.[7]
브로카의 환자들
루이 빅토르 르보르네 (탄)
레보르네는 브로카의 환자였습니다. 30살이 되던 해, 그는 거의 어떤 단어나 구절도 만들어낼 수 없었습니다.[45] 그는 반복적으로 "시간"이라는 단어만을 만들어낼 수 있었습니다. 그가 죽은 후, 그의 왼쪽 전두엽 표면에서 신경성 매독 병변이 발견되었습니다.
레롱
렐롱은 브로카의 또 다른 환자였습니다. 그는 또한 생산적인 말솜씨가 떨어졌습니다. 그는 '예', '아니오', '셋', '언제나', '엘로' (자신의 이름을 잘못 발음한) 다섯 단어만 말할 수 있었습니다. 레롱의 부검 과정에서 측두엽 내에 병변이 발견되었습니다. 브로카의 이전 환자인 레보르네는 전두엽과 같은 부위에 병변이 있었습니다. 이 두 가지 사례로 인해 브로카는 연설이 이 특정 영역에 국한되어 있다고 생각하게 되었습니다.[7]
MRI 소견
고해상도 MRI로 브로카의 두 역사적인 환자의 뇌를 조사한 결과 몇 가지 흥미로운 결과가 나왔습니다. 첫째, MRI 결과는 브로카 부위 외에 다른 부위도 환자들의 생산적인 말소리 감소에 기여했을 수 있음을 시사합니다. 이 발견은 브로카 부위의 병변만으로도 일시적인 언어 장애를 일으킬 수 있지만 심각한 언어 정지를 초래하지는 않는다는 사실을 밝혀냈기 때문에 의미가 있습니다. 따라서 브로카가 생산적인 말이 없는 것으로 표시한 실어증도 다른 지역의 병변의 영향을 받았을 가능성이 있습니다.[citation needed] 또 다른 발견은 한때 브로카에 의해 연설에 중요한 것으로 여겨졌던 그 지역이 현재 브로카의 지역으로 알려진 것과 정확히 같은 지역이 아니라는 것입니다. 이 연구는 언어와 인지가 한 때 생각했던 것보다 훨씬 더 복잡하고 다양한 뇌 영역의 네트워크를 포함한다는 주장을 뒷받침하는 추가 증거를 제공합니다.[46]
언어의 진화
인간의 언어 기원을 다루는 만족스러운 이론의 추구는 많은 진화론적인 "모델"을 고려하게 되었습니다. 이 모델들은 현대 언어가 어떻게 진화했을지 보여주려고 시도하는데, 이 이론들 중 많은 것들의 공통적인 특징은 음성적 의사소통이 제스처를 통해 훨씬 더 지배적인 의사소통 방식을 보완하기 위해 처음에 사용되었다는 생각입니다. 인간의 언어는 "손/입의 목표 지향적인 행동 표현을 기반으로 한 하위 영장류에 이미 존재하는 암묵적인 의사소통 시스템의 진화적 개선"으로 진화했을 수 있습니다.[19]
"손/입 목표 지향 행동 표현"은 "제스처 커뮤니케이션", "제스처 언어" 또는 "바디 언어를 통한 의사소통"을 말하는 또 다른 방식입니다. 최근 사람들이 다른 사람들이 의미 있는 행동을 하는 것을 관찰할 때 브로카의 영역이 활성화된다는 발견은 이 아이디어를 뒷받침하는 증거입니다. 현대 브로카 영역의 전신은 타인의 움직임을 지적인 목적을 가진 의미 있는 행동으로 해석함으로써 몸짓을 추상적인 아이디어로 변환하는 데 관여하고 있다는 가설을 세웠습니다. 시간이 지남에 따라 일련의 움직임의 의도된 결과와 목적을 예측하는 능력은 결국 이 영역에 진정으로 추상적인 생각을 다룰 수 있는 능력을 제공했고, 따라서 (결국) 소리(단어)와 추상적인 의미를 연관시킬 수 있게 되었다고 주장합니다. 사람들이 손그림자를[19] 관찰할 때 전두 언어 영역이 활성화된다는 관찰은 제스처 인식을 목적으로 진화한 기존 신경 기질에서 인간 언어가 진화했을 수 있다는 추가 증거입니다.[47] 따라서 이 연구는 브로카의 영역이 몸짓 언어와 몸짓을 해석하는 것과 같은 방식으로 말소리를 조립하고 해독하는 "말하기 위한 운동 중심지"라고 주장합니다. 이 생각과 일치하는 것은 유인원과 인간의 공통 조상에서 운동 조절을 조절하는 신경 기질이 인지 및 언어 능력을 향상시키기 위해 변형되었을 가능성이 가장 높다는 것입니다.[23] 미국 수화와 영어 사용자들에 대한 연구는 인간의 뇌가 훨씬 이전에 더 기본적인 기능을 수행하도록 진화한 시스템을 모집했음을 시사합니다; 저자들에 따르면, 이러한 다양한 뇌 회로들은 언어를 만드는 데 함께 일하기 위해 사용되었습니다.[48]
또 다른 최근의 발견은 침팬지의 의사소통적인 수동 제스처와 목소리 신호를 생산하는 동안 피질하부와 신피질하부에서 활성화의 중요한 영역을 보여주었습니다.[49] 게다가, 침팬지가 인간과 의사소통하기 위해 의도적으로 목소리 신호뿐만 아니라 수동 제스처를 만들어낸다는 것을 나타내는 데이터는 인간 언어의 전조가 행동적인 수준과 신경 해부학적인 수준 모두에 존재한다는 것을 암시합니다. 보다 최근에, 마모셋에서 활동 의존적인 유전자 발현의 신피질 분포는 인간의 브로카 영역을 구성하고 마카크의 종별 발성 및 구강 얼굴 제어의 청각 처리와 관련이 있는 복측 전전두엽 피질이 직접적인 증거를 제공했습니다. 신세계 원숭이의 목소리 출력 중에 참여합니다.[50][51] 이러한 연구 결과는 발성과 관련된 신피질 회로의 기원을 구세계와 신세계 원숭이 계통이 갈라졌던 적어도 3500만 년 전으로 추정하고 있습니다.
추가 영상
-
 브로카 지역(빨간색으로 표시됨). 애니매이션.
브로카 지역(빨간색으로 표시됨). 애니매이션. -
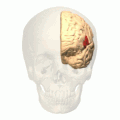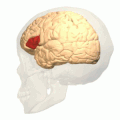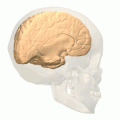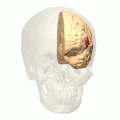
 회색으로 강조 표시된 브로카 지역의 대략적인 위치.
회색으로 강조 표시된 브로카 지역의 대략적인 위치. -

- 인간의 뇌 해부 비디오(24초). 하전두회에서 브로카 부위의 위치를 보여줍니다.
-
 브로카 지역의 시상 단면
브로카 지역의 시상 단면 -
 브로카 지역의 관상면
브로카 지역의 관상면 -
 브로카 지역의 횡단면
브로카 지역의 횡단면






참고 항목
참고문헌
- ^ "Broca's area". The American Heritage Dictionary of the English Language (5th ed.). HarperCollins. Retrieved 8 September 2019.
- ^ "Broca's area". Lexico UK English Dictionary. Oxford University Press. Archived from the original on 2022-08-26.
- ^ "Broca's area". Merriam-Webster.com Dictionary. Retrieved 8 September 2019.
- ^ "Broca's area". Collins English Dictionary. HarperCollins. Retrieved 8 September 2019.
- ^ Cantalupo C, Hopkins WD (November 2001). "Asymmetric Broca's area in great apes". Nature. 414 (6863): 505. Bibcode:2001Natur.414..505C. doi:10.1038/35107134. PMC 2043144. PMID 11734839.
- ^ Kennison, Shelia (2013). Introduction to language development. Los Angeles: Sage.[페이지 필요]
- ^ a b c d e Dronkers NF, Plaisant O, Iba-Zizen MT, Cabanis EA (May 2007). "Paul Broca's historic cases: high resolution MR imaging of the brains of Leborgne and Lelong". Brain. 130 (Pt 5): 1432–41. doi:10.1093/brain/awm042. PMID 17405763.
- ^ Hagoort, P. (2014). "Nodes and networks in the neural architecture for language: Broca's region and beyond" (PDF). Current Opinion in Neurobiology. 28: 136–141. doi:10.1016/j.conb.2014.07.013. hdl:11858/00-001M-0000-0019-FEFE-C. PMID 25062474. S2CID 16858289. Archived (PDF) from the original on 2022-10-09.
- ^ a b Plaza M, Gatignol P, Leroy M, Duffau H (August 2009). "Speaking without Broca's area after tumor resection". Neurocase. 15 (4): 294–310. doi:10.1080/13554790902729473. PMID 19274574. S2CID 16683208.
- ^ Grodzinsky Y, Santi A (December 2008). "The battle for Broca's region". Trends in Cognitive Sciences. 12 (12): 474–80. doi:10.1016/j.tics.2008.09.001. PMID 18930695. S2CID 13023258.
- ^ a b c d Skipper JI, Goldin-Meadow S, Nusbaum HC, Small SL (June 2007). "Speech-associated gestures, Broca's area, and the human mirror system". Brain and Language. 101 (3): 260–77. doi:10.1016/j.bandl.2007.02.008. PMC 2703472. PMID 17533001.
- ^ Schlaepfer TE, Harris GJ, Tien AY, Peng L, Lee S, Pearlson GD (1995). "Structural differences in the cerebral cortex of healthy female and male subjects: a magnetic resonance imaging study". Psychiatry Res. 61 (3): 129–35. doi:10.1016/0925-4927(95)02634-a. PMID 8545497. S2CID 25425465.
- ^ Caplan D (May 2006). "Why is Broca's area involved in syntax?". Cortex; A Journal Devoted to the Study of the Nervous System and Behavior. 42 (4): 469–71. doi:10.1016/S0010-9452(08)70379-4. PMID 16881251. S2CID 4480694.
- ^ Grewe T, Bornkessel I, Zysset S, Wiese R, von Cramon DY, Schlesewsky M (November 2005). "The emergence of the unmarked: a new perspective on the language-specific function of Broca's area". Human Brain Mapping. 26 (3): 178–90. doi:10.1002/hbm.20154. PMC 6871720. PMID 15929098.
- ^ Rodd JM, Davis MH, Johnsrude IS (August 2005). "The neural mechanisms of speech comprehension: fMRI studies of semantic ambiguity". Cerebral Cortex. 15 (8): 1261–9. doi:10.1093/cercor/bhi009. PMID 15635062.
- ^ Devlin JT, Matthews PM, Rushworth MF (January 2003). "Semantic processing in the left inferior prefrontal cortex: a combined functional magnetic resonance imaging and transcranial magnetic stimulation study". Journal of Cognitive Neuroscience. 15 (1): 71–84. CiteSeerX 10.1.1.329.8485. doi:10.1162/089892903321107837. PMID 12590844. S2CID 7570128.
- ^ Nixon P, Lazarova J, Hodinott-Hill I, Gough P, Passingham R (March 2004). "The inferior frontal gyrus and phonological processing: an investigation using rTMS" (PDF). Journal of Cognitive Neuroscience. 16 (2): 289–300. doi:10.1162/089892904322984571. PMID 15068598. S2CID 1162060.
- ^ Gough PM, Nobre AC, Devlin JT (August 2005). "Dissociating linguistic processes in the left inferior frontal cortex with transcranial magnetic stimulation". The Journal of Neuroscience. 25 (35): 8010–6. doi:10.1523/JNEUROSCI.2307-05.2005. PMC 1403818. PMID 16135758.
- ^ a b c d Fadiga L, Craighero L, Destro MF, Finos L, Cotillon-Williams N, Smith AT, Castiello U (2006). "Language in shadow". Social Neuroscience. 1 (2): 77–89. doi:10.1080/17470910600976430. PMID 18633777. S2CID 20322.
- ^ Fadiga L, Craighero L (May 2006). "Hand actions and speech representation in Broca's area". Cortex; A Journal Devoted to the Study of the Nervous System and Behavior. 42 (4): 486–90. doi:10.1016/S0010-9452(08)70383-6. PMID 16881255. S2CID 2463077.
- ^ Gentilucci M, Bernardis P, Crisi G, Dalla Volta R (July 2006). "Repetitive transcranial magnetic stimulation of Broca's area affects verbal responses to gesture observation". Journal of Cognitive Neuroscience. 18 (7): 1059–74. doi:10.1162/jocn.2006.18.7.1059. PMID 16839281. S2CID 18159912.
- ^ a b Carlson, N. (2013). "Human Communication". Physiology of Behavior (11th ed.). Boston: Allyn and Bacon.
- ^ a b Lieberman P (2002). "On the nature and evolution of the neural bases of human language". American Journal of Physical Anthropology. Suppl 35: 36–62. doi:10.1002/ajpa.10171. PMID 12653308.
- ^ "Aphasia: Signs & Symptoms".
- ^ Maguire et al. 1994[full citation needed]
- ^ Maguire EA, Frackowiak RS, Frith CD (September 1997). "Recalling routes around london: activation of the right hippocampus in taxi drivers". The Journal of Neuroscience. 17 (18): 7103–10. doi:10.1523/JNEUROSCI.17-18-07103.1997. PMC 6573257. PMID 9278544.
- ^ a b c "What is Aphasia". Atlanta Aphasia Association. 2006. Archived from the original on 2008-12-23. Retrieved 2008-12-01.
- ^ Metz-Lutz MN, Dahl E (September 1984). "Analysis of word comprehension in a case of pure word deafness". Brain and Language. 23 (1): 13–25. doi:10.1016/0093-934X(84)90002-6. PMID 6478188. S2CID 39218546.
- ^ a b Slevc LR, Martin RC, Hamilton AC, Joanisse MF (January 2011). "Speech perception, rapid temporal processing, and the left hemisphere: a case study of unilateral pure word deafness". Neuropsychologia. 49 (2): 216–30. doi:10.1016/j.neuropsychologia.2010.11.009. PMC 3031136. PMID 21093464.
- ^ Poeppel, David (2001). "Pure word deafness and the bilateral processing of the speech code". Cognitive Science. 25 (5): 679–93. doi:10.1207/s15516709cog2505_3.
- ^ "The National Aphasia Foundation". Archived from the original on January 22, 2011. Retrieved January 15, 2011.
- ^ Friedenberg J, Silverman G (2006). "Evaluating Techniques for the Study of Brain Damage". Cognitive science: an introduction to the study of mind. pp. 165–6. ISBN 978-1-4129-2568-6.
- ^ Mohr JP, Pessin MS, Finkelstein S, Funkenstein HH, Duncan GW, Davis KR (April 1978). "Broca aphasia: pathologic and clinical". Neurology. 28 (4): 311–24. doi:10.1212/WNL.28.4.311. PMID 565019. S2CID 34920053.
- ^ Kaan E, Swaab TY (August 2002). "The brain circuitry of syntactic comprehension". Trends in Cognitive Sciences. 6 (8): 350–356. doi:10.1016/S1364-6613(02)01947-2. PMID 12140086. S2CID 18668619.
- ^ Dronkers NF, Shapiro JK, Redfern B, Knight RT (1992). "The role of Broca's area in Broca's aphasia". Journal of Clinical and Experimental Neuropsychology. 14: 52–3.
- ^ Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR (October 1996). "Brain activation modulated by sentence comprehension". Science. 274 (5284): 114–6. Bibcode:1996Sci...274..114J. doi:10.1126/science.274.5284.114. PMID 8810246. S2CID 30517695.
- ^ Friedman L, Kenny JT, Wise AL, Wu D, Stuve TA, Miller DA, Jesberger JA, Lewin JS (September 1998). "Brain activation during silent word generation evaluated with functional MRI". Brain and Language. 64 (2): 231–56. doi:10.1006/brln.1998.1953. PMID 9710491. S2CID 46640048.
- ^ Binkofski F, Amunts K, Stephan KM, Posse S, Schormann T, Freund HJ, Zilles K, Seitz RJ (December 2000). "Broca's region subserves imagery of motion: a combined cytoarchitectonic and fMRI study" (PDF). Human Brain Mapping. 11 (4): 273–85. doi:10.1002/1097-0193(200012)11:4<273::AID-HBM40>3.0.CO;2-0. PMC 6872088. PMID 11144756.
- ^ Talairach J, Tournoux P (1988). Co-planar stereotaxic atlas of the human brain. New York: Thieme Medical.[페이지 필요]
- ^ Dronkers NF, Wilkins DP, Van Valin RD, Redfern BB, Jaeger JJ (2004). "Lesion analysis of the brain areas involved in language comprehension". Cognition. 92 (1–2): 145–77. doi:10.1016/j.cognition.2003.11.002. hdl:11858/00-001M-0000-0012-6912-A. PMID 15037129. S2CID 10919645.
- ^ D'Esposito M, Postle BR, Ballard D, Lease J (October 1999). "Maintenance versus manipulation of information held in working memory: an event-related fMRI study". Brain and Cognition. 41 (1): 66–86. doi:10.1006/brcg.1999.1096. PMID 10536086. S2CID 14336072.
- ^ Friederici AD (February 2002). "Towards a neural basis of auditory sentence processing". Trends in Cognitive Sciences. 6 (2): 78–84. doi:10.1016/S1364-6613(00)01839-8. PMID 15866191.
- ^ Burton MW, Small SL, Blumstein SE (July 2000). "The role of segmentation in phonological processing: an fMRI investigation". Journal of Cognitive Neuroscience. 12 (4): 679–90. doi:10.1162/089892900562309. PMID 10936919. S2CID 685383.
- ^ Flinker A, Chang EF, Barbaro NM, Berger MS, Knight RT (June 2011). "Sub-centimeter language organization in the human temporal lobe". Brain and Language. 117 (3): 103–9. doi:10.1016/j.bandl.2010.09.009. PMC 3025271. PMID 20961611.
- ^ Konnikova, Maria. "The man who couldn t speak and how he revolutionized psychology". Scientific American Blog Network. Archived from the original on 2016-09-14. Retrieved 2017-05-03.
- ^ "Anatomy of Speech & Language UCSF Memory and Aging Center". memory.ucsf.edu. Archived from the original on 2017-05-03. Retrieved 2017-05-03.
- ^ Corballis MC (April 2003). "From mouth to hand: gesture, speech, and the evolution of right-handedness". The Behavioral and Brain Sciences. 26 (2): 199–208, discussion 208–60. doi:10.1017/S0140525X03000062. PMID 14621511. S2CID 21861033.
- ^ Newman AJ, Supalla T, Hauser P, Newport EL, Bavelier D (April 2010). "Dissociating neural subsystems for grammar by contrasting word order and inflection". Proceedings of the National Academy of Sciences of the United States of America. 107 (16): 7539–44. Bibcode:2010PNAS..107.7539N. doi:10.1073/pnas.1003174107. PMC 2867749. PMID 20368422.
- "Sign language study shows multiple brain regions wired for language". ScienceDaily (Press release). April 30, 2010.
- ^ Taglialatela JP, Russell JL, Schaeffer JA, Hopkins WD (March 2008). "Communicative signaling activates 'Broca's' homolog in chimpanzees". Current Biology. 18 (5): 343–8. doi:10.1016/j.cub.2008.01.049. PMC 2665181. PMID 18308569.
- ^ Simões CS, Vianney PV, de Moura MM, Freire MA, Mello LE, Sameshima K, Araújo JF, Nicolelis MA, Mello CV, Ribeiro S (2010). "Activation of frontal neocortical areas by vocal production in marmosets". Frontiers in Integrative Neuroscience. 4. doi:10.3389/fnint.2010.00123. PMC 2955454. PMID 20953246.
- ^ Miller CT, Dimauro A, Pistorio A, Hendry S, Wang X (2010). "Vocalization Induced CFos Expression in Marmoset Cortex". Frontiers in Integrative Neuroscience. 4: 128. doi:10.3389/fnint.2010.00128. PMC 3004388. PMID 21179582.
외부 링크
- "폴 브로카의 관절 언어를 지배하는 뇌 영역의 발견", 브로카의 1861년 기사를 분석한 BibNum [영어 버전은 'a télacharger' 클릭].

